
�ҹ�ÿ��ũ���Ӳ�Σ����������9.3�ڹ���,�ɴ���ɵľ�����ʧ��2,200��Ԫ,���ݼ���ʹ������˾����ʧ��Ȼ��,���ݼ�����������������,�ѳ�Ϊ��вȫ����ʳ��ȫ�Ĺؼ�����֮һ����ֹ2022��11��,ȫ�����ѱ���515�ֳ��ݼ������Ӳ�������,����267�ֲַ���72������,�漰97������ҹ����������Ӳݿ���Σ����Ϊ���ص�5������֮һ,���п����Ӳ�������С�����ˮ�������ͬ���ͻ��ֵĿ����ݼ��Ӳ������ͼ1��ͼ1��Ϊ������ģʽ���ֵĿ����ݼ��Ӳ�����,��ͼ�п��Կ���,��ͬ�б��Ӳݿ�����������������,��������ģʽΪ���������ø���Ӳݿ���������ࣻ��ͼ1�ҿ��Կ���,ݬȥ�������ֲ�����������һ����ݼ�,����Dzݸ�좡���Щ�����Ӳ�Σ������,��������,�����ݼ��·��Ӱб�ķ��������ڻ��⿹���Ӳ���һ�������⡣Ϊ��,����������Ȫ��̷Ч�ɵ��ܽ���ݼ����Ӱб����ͺ��ܵĻ�����,�ع��˽�30�������ֵij��ݼ����Ӱб�,���ص���ܷ��Ӱб���������ܡ����ݼ����û��ơ����ݼ����÷�Χ�Լ��б���ӽṹ���о���չ��ͬʱ���ܵ�ǰ�°б귢�ֵ��������б귢�ֵ��¼����Լ���δ�����ݼ��б귢�ֵ�չ����
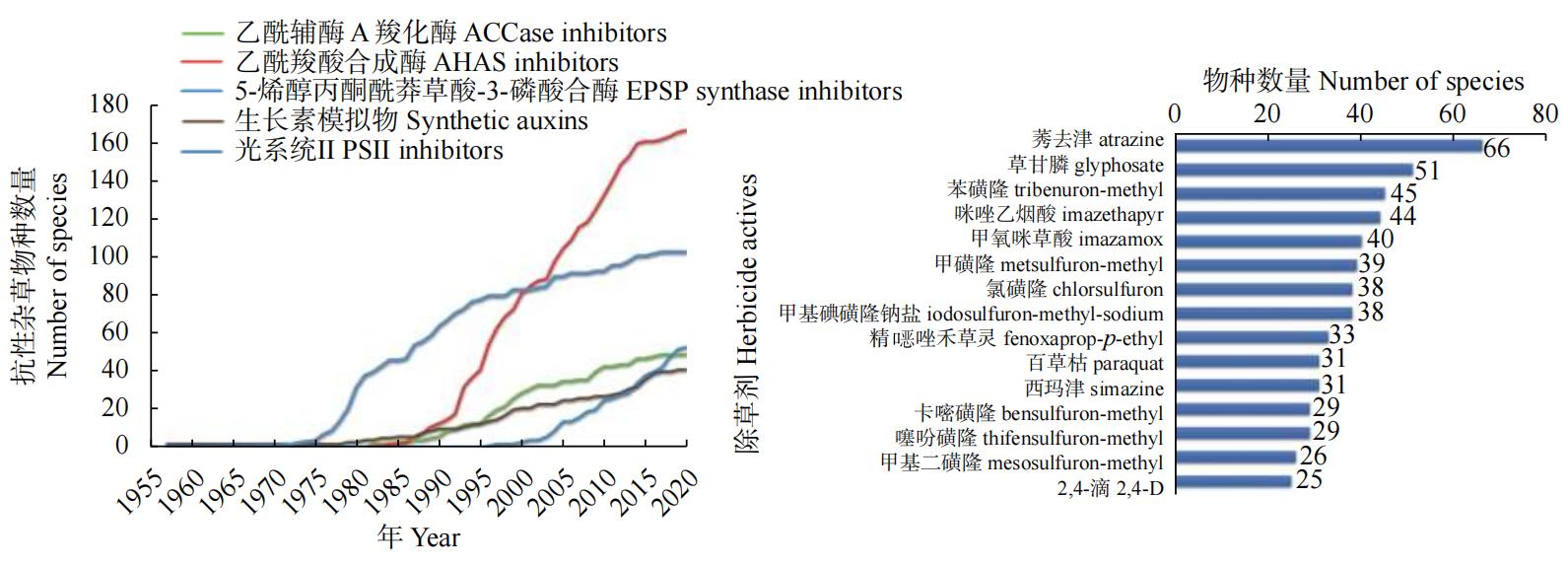
ͼ1 �����Գ��ݼ������������ðе�Ŀ���ͳ��
1 ��30��ij��ݼ����Ӱб����
���ݼ������ðб��Ϊֲ���������л;�еĸ���ø,���ż�������Ƕ���������(ADP)��������������(ATP)�������ữ���̵Ĵ�ø,ԭ߲��ԭ����ø������Ѫ���غ�Ҷ���صĺϳ�,��Щø��Ϊֲ����������ø,��ȱʧ�������Щø�ĺϳ�;��,���ᵼ��ֲ����ֲ�֢ͬ״�����������ճ��ݼ����û��ƵIJ�ͬ,�ɽ����ݼ���Ϊ������������Ƽ��������������Ƽ�������ϳ����Ƽ�����ά�غϳ���ء��������Ƽ���������ϳ���غ�����7��(��1)�������ص���ܳ��ݼ����û��ƶ�Ӧ�ķ��Ӱб���������ܡ����ݼ������û��ƺʹ�����Ʒ��(���ζ����Ӧ�÷�Χ)��
��1 ���ݼ��б�ķ��༰������Գ��ݼ�Ʒ��


2 ���ݼ����Ӱб���������ܺ���ά�ṹ�о�
������ӳ��ݼ�ʩ�ú�,Ӱ���Ӳݹ�����á��������á�����ϳɡ���ά�غϳɡ��������ơ�������ϳ���غ�����7�����,��ϸ���ܳ��ݼ����Ӱб���������ܡ����ݼ�ʩ�ö���ʩ�÷�Χ,Ϊ�˽����г��ݼ����Ӱб��Լ�ͬһ���Ӱб겻ͬ�������ռ���ԵIJ�����,��ÿ���б겻ͬ�����ĵ�����������Ӧ��A����ά����ͼ��
2.1 ����������Ƽ�
2.1.1 ��ϵͳI���Ӵ���
�����������ɫֲ��(����ijЩϸ��)������ת��Ϊ���������������л�����������̡���������ַ�Ϊ�������̣���ϵͳ���ϵͳ��ϵͳ��϶�ֲ���������Ĥ����Ĥ��,���ɵ����ǻ���ɵ�ɫ�ص�������,����Ҫ����Ϊ���ڹ�ϵ��Ӵ�������,�����Ӵ�������Ĥ��ת�Ƶ�Ĥ������������,������������--��ø��(NADP)��ԭø������,��NADP+��ԭΪ��ԭ��ø��(NADPH)�����ƹ�ϵͳ��ij��ݼ������ֲ��Ĺ���Ӵ���,�ƻ���ԭ��ø����γ�,�������ֲ����й�����á��ٲݿ�(paraquat)��������ݼ��Ĵ���,�����û���Ϊ����һ��������,ֲ����Ĥ�����ĵ�����ٲݿ����ӷ�Ӧ�������ɻ�,�����ɻ���ֲ�����ڵ�������ԭ,����������(ROS),����ROS�����ܹ���ϸ��Ĥ�IJ�����֬�������֬���ɻ�,����Ѹ�ٴ��ֽ�ϸ��Ĥ����֯,����ֲ������ϸ����֯����ʧˮ,���ֲ����ʧˮ�������ή������
2.1.2 ��ϵͳ���D1����
��ϵͳ����ֲ�ォ���Ӵ�ˮ��ת�Ƶ���������,����������D1����ϡ�D1�����ǹ�ϵͳ��Ӧ����һ����Ҫ���ǻ�����,�õ���Ϊ���ָ��������ṩ���λ��,��ԭ����ɷ���ʹ����й�,���D1���ױ��ƻ�,�ᵼ�¹�ϵ��Ӵ����������Ӱ���ϵͳ��Ӧ����,��Ӱ�������á����ƹ�ϵͳ��ij��ݼ���һ��������������D1����ҩ��,ͨ����D1�����,��ϵ��Ӵ�QAλ�㴫��QBλ��,���ƹ�����õķ�����ݬȥ��(atrazine)��������ݼ��Ĵ���,��Ҫ�������ס����ᡢ�����ȵص�ѿǰ��ѿ��ѧ���ݡ�ʩ��ݬȥ���,�Ӳ�ͨ����������ݬȥ�����ϴ���,ݬȥ���������������D1���ס���ݬȥ����D1����Ϻ�,��������������D1���Ľ�Ϻ͵��Ӵ�������,��Щ���Ӿͻ���ϸ��Ĥ�е���֬��Ӧ,�ƻ�ϸ��Ĥ,���յ����Ӳ�������
2.1.3 ���ⷬ�Ѻ���ȥ����ø
����ܲ��ز���ֲ��������,����������Ĥ�Ϲ��Ԫ���ؼ��ṹ����ɲ��֡����ⷬ�Ѻ���ȥ����ø(PDS)������ܲ��غϳ�;���е�һ������ø,���ⷬ�Ѻ���ͨ��PDS��������,�γ���-���ܲ���,��-���ܲ�������-���ܲ�������ø��������,�����γɷ��Ѻ��ء���ͪ(flurtamone)��������ݼ��Ĵ���,��ͨ�������Ӳ��е�PDS,�谭����ܲ��صĺϳɶ��ﵽ����Ч����
����ѡȡ�˲��ܷ���Pantoea ananatis(PDB��ţ�4DGK,�ֱ��ʣ�2.35 Å (1 Å=0.1 nm))��ˮ��Oryza sativa L.(PDB��ţ�5MOG,�ֱ��ʣ�2.77 Å)PDSA���������ṹ����ͼ(ͼ2),����pymol��������,�ó�������ƫ��(RMSD)ԼΪ10.034 Å,ˮ������(��ɫ)�ṹ�Ͳ��ܷ�������(��ɫ)�ṹ��ͬ�㣺���߶���3���ṹ��,�����۵����ƣ��䲻ͬ�㣺���ܷ����Ļ��Կ�ǻ��ˮ�����Դ�
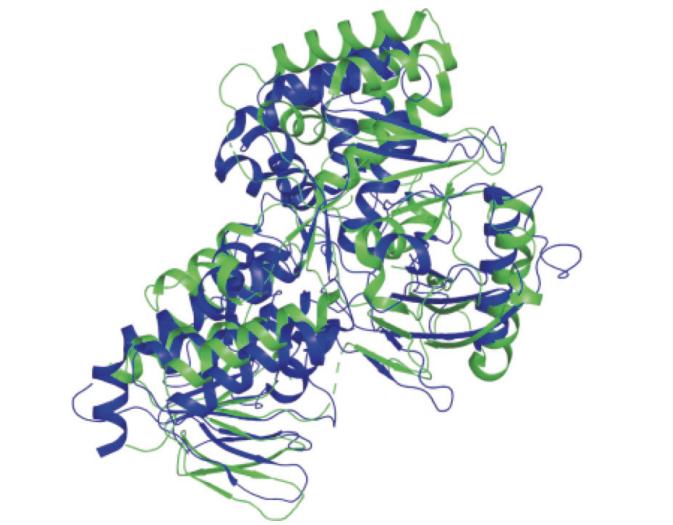
ͼ2 ˮ���Ͳ��ܷ�������A����ά�ṹ����ͼ
2.1.4 ���Ѻ��ػ���ø
���Ѻ��ػ���ø�������Ѻ�����-����ø�ͷ��Ѻ�����-����ø,���߶�������ܲ�������ϳ�;����ø�����Ʒ��Ѻ��ػ���ø�ij��ݼ�������ɱ��ǿ(amitrole),�ó��ݼ����ڿ���һ�����Ӳݺ�ˮ��(������������ʳ�������Ӳݵķ���)��
2.1.5 ԭ߲��ԭ����ø
ԭ߲��ԭ����ø(PPO)��ֲ����������������ϳ�;�������һ��ø��PPOͨ����ԭ߲��ԭ������ԭ߲����,ԭ߲������Ҷ���غ�Ѫ���غϳ�;�������ʡ�����PPO�ij��ݼ���Ҫ���û����ǣ����ݼ�����PPO,���ԭ߲��ԭ���Ļ���,����ԭ߲��ԭ����й��ϸ������,��ϸ������Ѹ�ٱ�������ԭ߲����,��ϸ�����е�ԭ߲�������ܱ�����,����ֲ���������ܵ�Ӱ�졣����PPO�ij��ݼ�������˫�����(pyraclonil),˫������Ǵ�ɱ�ͳ��ݼ�,ͨ������PPO����ҩЧ����,����Ч���Ƹ����Ӳ�,��ݡ���Ҷ�Ӳݺ�ɯ���Լ��ͻ���������ݼ����Ӳݵȡ�
����ѡȡ�˿ݲ�ѿ�߸˾�Bacillus subtilis(PDB��ţ�3I6D,�ֱ��ʣ�2.90 Å)���̲�Nicotiana tabacum L.(PDB��ţ�1SEZ,�ֱ��ʣ�2.90 Å)����ɫճ���Myxococcus xanthus(PDB��ţ�2IVD,�ֱ��ʣ�2.30 Å)����Homo sapiens(PDB��ţ�3NKS,�ֱ��ʣ�1.90 Å)��PPO A���������ṹ����ͼ,��ͼ3����ͼ3��֪�ݲ�ѿ�߸˾�����(��ɫ)���̲ݵ���(��ɫ)���ϳ̶ȽϺ�,��ɫճ���(��ɫ)����(�ػ�ɫ)�ĵ��ṹ��ݲ�ѿ�߸˾����̲ݵ������ƶȲ���,�������Կ�ǻ�Դ�
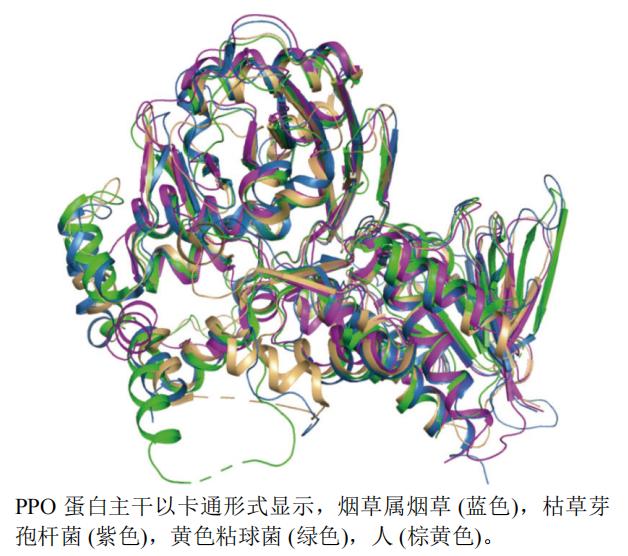
ͼ3 PPO��ͬ����A����ά�ṹ����
2.2 �����������Ƽ�--��ż������
��ż����ָ������������������ữ��ż����ƻ�������ż����(���ƽ�ż��)������ֲ���������ữ��λ��,��ADP����ATP�ķ�Ӧ�ܵ�����,����ADPŨ������,��Ȼ������ǿֲ���������,ȴ��������ATP������ֲ����������������,���յ���ֲ���л��������ݲ�¡(neburon)�ͻ���¡(siduron)���������龥���������������ữ�Ľ�ż������
2.3 ����ϳ����Ƽ�
2.3.1 ������øA�Ȼ�ø
ֲ��֬��������֬�Լ���֬����Ҫ��ɲ���,����Ҫ����Դ�����Լ��źŴ������ӡ�������øA�Ȼ�ø(ACCase)����������øA�ĺϳ�,��������øA������������֬����ϳɡ��������(cyhalofop-butyl)�Ƿ�������������������Ψһ��ˮ����ȫ�Ըߵij��ݼ�,ͨ�������Ӳ�ACCase�Ļ���,ʹ֬����ϳ�ֹͣ,ϸ�������ͷ��Ѳ�����������,���յ����Ӳ�����,��������������л����˲�Ҳ�нϺõķ���Ч����
����ѡȡ�˴˾�Escherichia coli(PDB��ţ�2F9Y,�ֱ��ʣ�3.20 Å)������ë�Ǿ�����DSM 1495(Chaetomium thermophilum var. thermophilum DSM 1495)(PDB��ţ�5I6I,�ֱ��ʣ�8.40 Å)����ƽ�ĸSaccharomyces cerevisiae(PDB��ţ�5CSA,�ֱ��ʣ�3.00 Å)��ACCase A���������ṹ����ͼ(ͼ4),����pymol��������,��ͼ�п��Կ�����3�൰�����ƶȲ���,����ë�Ǿ�����DSM1495(�ۺ�ɫ)����ƽ�ĸ����(��ɫ)�ṹ�ȴ˾�����(��ɫ)�ṹ��
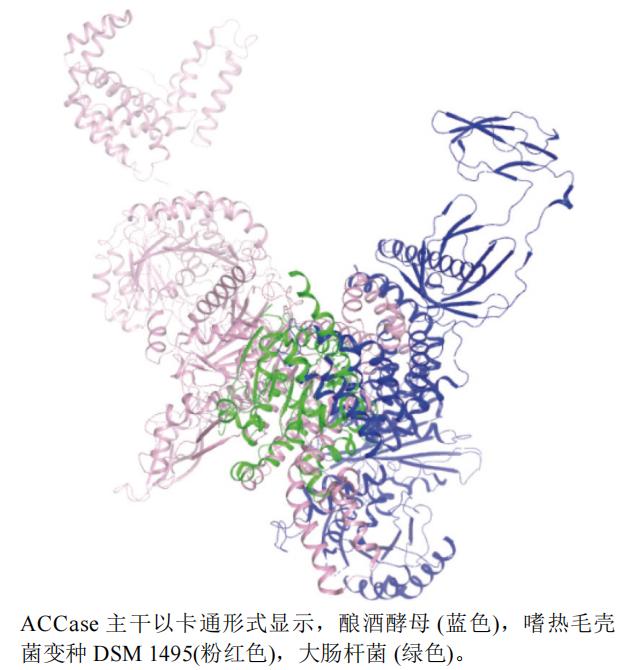
ͼ4 ACCase��ͬ����A����ά����ͼ
2.3.2 ֬��������ø
֬��������ø(FAT)��֬����ϳɹ����п�������,��ʹ֬������������嵰���ͷŵ�ø������FAT�ij��ݼ���ͨ����FAT���,����֬������ͷš���������(cinmethylin)������FAT����,��Ϊ��ǰ���ݼ�,�ѱ�����Ĵ����ǹ�����ֲ,���ڿ���һ��������ݡ����������(methiozolin)��һ�����ͳ��ݼ�,���ڿ��Ƽ���ů�����伾��ƺ���е�һ�������ݡ�
2.3.3 ������������ø
������֬��β����ɲ����Ƕ������Ѵ�,����ͨ��������������ø(SPS)�Ļ��Ի�õ�,SPS��������ϩ�����������μӳ��γ���Ҷ�������ᡣSPS��2020�걻Kahlau��֤ʵΪ���ݼ�������(aclonifen)�����ðб�,��һ��ȫ�µij��ݼ�����ģʽ��SPS������,�������Ӳ�����������ˮƽ�����½�,���յ����Ӳݱ�Ư�ס�
2.3.4 �����������ת��ø
�����������ת��ø(HST)������������ϳ�;���ж��ǻ�������ͪ��˫����ø������ø��HST������������ϩ���������γ�2-��-6-�����-1,4����୴�,��������������ϳ��еĵ�һ���м��塣Shin����2018��֤ʵ,���ݼ�����າ���(cyclopyrimorate)�İ�λ����HST,HST����Ϊ��һ�����͵���ҵ���ݼ����Ӱбꡣ
2.3.5 ��������ϳ�ø
���������ø,Ҳ��Ϊ�����ǻ����ø(AHAS)��AHAS��֧������������ϳ�;���е�һ���ؼ�ø,֧����������������ᡢ����������Ӱ���,��ֲ��ȱ����3�ְ�����,��Ӱ�쵰���ʺϳ�,����ϸ������,�����Ӳ���֯ʧ�̡��ƻ�����������,����Ӳ������������ݼ�����Dzݰ�(penoxsulam)���ðе���AHAS,Ϊ�����ݼ�,����ˮ�����Ӳݷ��Ρ�����Dzݰ������Ӳ���AHAS�Ľ�ϲ�λ�����Խ��,�Ӷ�Ӱ���Ӳ����ڰ�����ĺϳ�,�����Ӳ�����,�Ӳ���������ʧ�̶�������
����ѡȡ�˹���ľùTrichoderma harzianum(PDB��ţ�7EGV,�ֱ��ʣ�2.54 Å)�������ײ��Ͼ�Klebsiella pneumoniae(PDB��ţ�1OZG,�ֱ棺2.30 Å)���������۾�MSB8 Thermotoga maritima MSB8(PDB��ţ�2FGC,�ֱ��ʣ�2.30 Å)�Ϳݲ�ѿ�߸˾�B. subtilis(PDB��ţ�4RJI,�ֱ��ʣ�3.20 Å)��AHASA��]�������ṹ����ͼ,��ͼ5��������ͼ�Կݲ�ѿ�߸˾�����(��ɫ)Ϊ������е���,���Կ���,���˺������۾�����(��ɫ)�ṹ��������ͬ,���൰�ṹ���ơ�
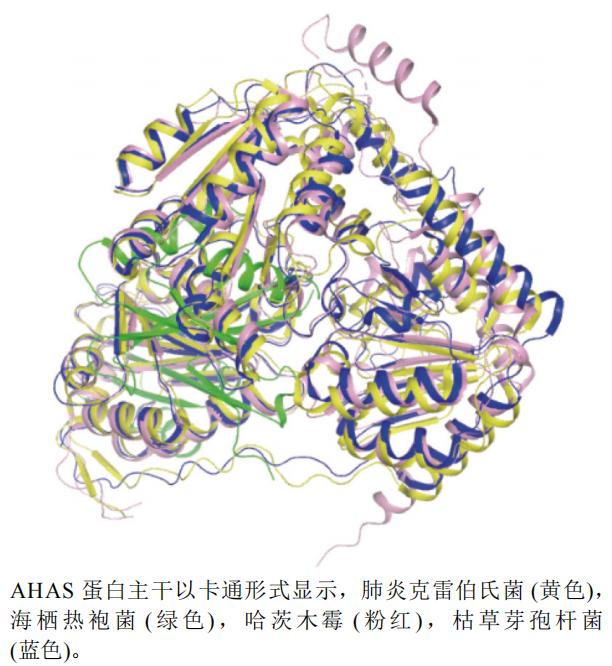
ͼ5 AHAS��ͬ����A����ά����ͼ
2.3.6 ���ǻ�����ˮø
���ǻ�����ˮø(DHAD)��֧������������ϳ�;����һ�ֳ���ø,�ڲ�ͬ��ֲ�������и߶ȱ��ء��о�֤��,DHAD��һ�����õij��ݼ����Ӱб�,������DHAD�ij��ݼ�����Բ��鶯�����Ӱ��,��Ϊ���鶯�����ڲ�������֧������������ϳ�;�������ݴ�ѧ��ɼ����Уһ������С��ʹ�ÿ��Ի���Ϊ����Ļ������ھ���,��������Ȼ�����춬����(aspterric acid)������������DHAD,�Ӷ�������ֲ�����������á�
����ѡȡ�˼�����Synechocystis sp.(PDB��ţ�6NTE,�ֱ��ʣ�2.33 Å)�����Ͻ�Arabidopsis thaliana(PDB��ţ�5YMO,�ֱ��ʣ�1.84 Å)��DHADA���������ṹ����ͼ(ͼ6),�ó�RMSDԼΪ1.123 Å,�������ߵ��ṹ����,��������(��ɫ)�Ļ��Կ�ǻ�����Ͻ�(��ɫ)�Ļ��Կ�ǻ��
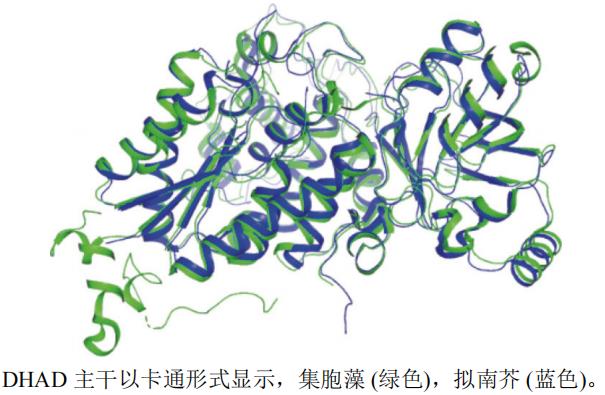
ͼ6 DHAD��ͬ����A����ά����ͼ
2.3.7 5-ϩ����ͪ��ç����-3-�����ø
5-ϩ����ͪ��ç����-3-�����ø(EPSPS)��Ҷ�����е�һ��ø,����ç����ϳ�;���ĵ�����2����Ӧ,�Ǻϳɷ����就�����Լ����ִ�����л����̵Ĺؼ�ø������EPSPS,�Ӷ�����ç���������ᡢ�Ұ��ἰɫ�����ת��,ʹ�����ʺϳ��ܵ�����,����ֲ���������ݸ��(glyphosate)�������Ӳݷ��Ӱб��EPSPS,�Ӳ�ͨ����Ҷ���պ���ֲ�����λ,�ɷ�������Ҷ��˫��Ҷ��һ���������������ݱ���ľ��40���ֲ�
����ѡȡ�˴˾�E. coli(PDB��ţ�1MI4,�ֱ��ʣ�1.70 Å)�����������Streptococcus pneumoniae(PDB��ţ�1RF4,�ֱ��ʣ�2.20 Å)�Ͱ�˿��ĸCandida albicans(PDB��ţ�7TBU,�ֱ��ʣ�1.85 Å)EPSPSA���������ṹ����ͼ(ͼ7)������pymol�����Դ˾�����(��ɫ)�ṹΪ������е��Ϻ���,��˿��ĸ����(�ۺ�ɫ)�ṹ���Կ�ǻ�ȴ˾��ͷ������������(��ɫ)���Կ�ǻ��
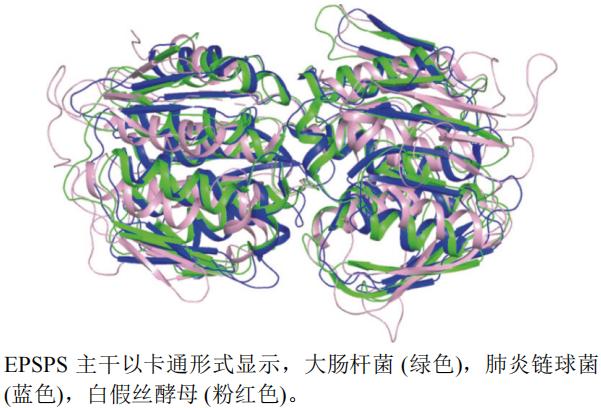
ͼ7 EPSPS��ͬ����A����ά����ͼ
2.3.8 ˿����-�հ��ᵰ������ø
˿����-�հ��ᵰ������ø���ṹ���ڶ��ֲ���ԭ�������и߶ȱ��ء�������������Ϊ˿����/�հ�������ø�ǰ�����(ˮ�ݼ׳����������Ӭ����Ȼ����)����������������ľù���İбꡣ˿����-�հ��ᵰ������ø�������(endotha)�İб�,������յ����ص���������,���ڹ���ˮ�������е��Ӳݡ�
����ѡȡ����ƽ�ĸSaccharomyces cerevisiae(PDB��ţ�3ICF,�ֱ��ʣ�2.30 Å)����H. sapiens(PDB��ţ�3H67,�ֱ��ʣ�1.65 Å)����˿��ĸCandida albicans(PDB��ţ�5JPE,�ֱ��ʣ�2.61 Å)����ϣ�Ͼ�������(Escherichia virus lambda)(PDB��ţ�1G5B,�ֱ��ʣ�2.15 Å)���ּ���Rattus norvegicus(PDB��ţ�4JA7,�ֱ��ʣ�2.00 Å)�����Ͻ�A. thaliana(PDB��ţ�5JJT,�ֱ��ʣ�2.10 Å)��˿����-�հ��ᵰ������øA���������ṹ����ͼ(ͼ8)������pymol����,����(��ɫ)�ĵ��ṹΪ������Ϻ���,��ϣ�Ͼ�������(��ɫ)�����Ͻ�(ī��ɫ)�ĵ��ṹ���Կ�ǻ�����������Կ�ǻ��
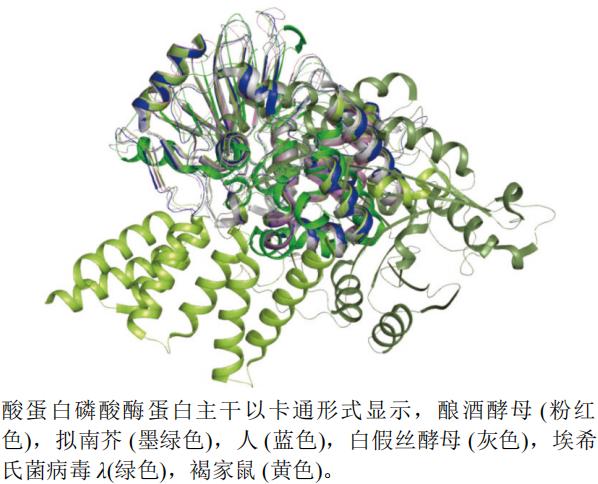
ͼ8 ˿����-�հ��ᵰ������ø��ͬ����A����ά����ͼ
2.3.9 �������������ˮø
�������������ˮøҲ��һ�����õij��ݼ��б�,�������ڲ������������������ˮø���������������ˮø��ֲ����������鰱������ϳɵ���Ҫ����,���鰱������ϳ�;���д��������������ת��Ϊ�������������������Ѿ�֤ʵ�������������ˮø���Ƽ����й��������ݻ���,3�������������������������������ˮø�Ļ���,ϸ����������ʾ��ǿ���������á�
2.3.10 ���DZ�����ͪ��˫����ø
���DZ�����ͪ��˫����ø(HPPD)�����������Ұ����л��������Ҫ��ø,�Ұ������Ұ��ᰱ��ת��ø�����������ɶ��ǻ�����ͪ��,�������IJ�����HPPD�ܹ������ǻ�����ͪ���ת��������ᡣ��ֲ������,������ܹ�����һ��ת�����������������ӡ���HPPD�Ļ��Ա�����,�Ӳ������Ұ���������л�������,�����Ӳ���������ܲ��ص�ȱ��,�Ӷ�ʹҶ���ع��������ü���,Ӱ��ֲ��Ĺ������,����ʹ�Ӳݰ������������Dz�ͪ(mesotrione)Ϊ���ص���������ݼ�,��������һ������Ҷ�ӲݺͲ��ֺ̱����Ӳ�(�����顢�Ȳˡ���ޤ���ݡ����Ƶ�)�нϺõķ���Ч��,�������Ȳ˺�һЩ�̱����Ӳݷ���Ч���ϲ
����ѡȡ������Zea mays L.(PDB��ţ�1SP8,�ֱ��ʣ�2.00 Å)��ӫ��ٵ�����Pseudomonas fluorescens(PDB��ţ�1CJX,�ֱ��ʣ�2.40 Å)����ά��ù��Streptomyces avermitilis(PDB��ţ�1T47,�ֱ��ʣ�2.50 Å)����H. sapiens(PDB��ţ�6ISD,�ֱ��ʣ�2.40 Å)�����Ͻ�A. thaliana(PDB��ţ�1SP9,�ֱ��ʣ�2.13 Å)��HPPDA������������ͼ(ͼ9)������pymol������ӫ��ٵ���������(��ɫ)Ϊ������е��Ϻ���,����5����������һ������������á�
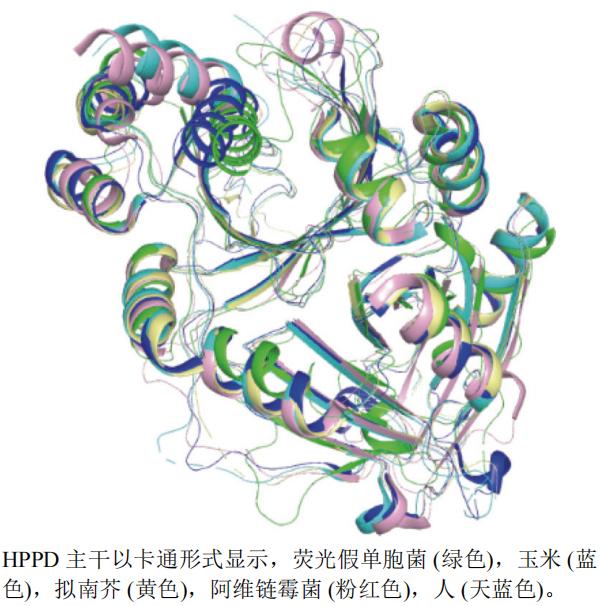
ͼ9 HPPD��ͬ����A����ά����ͼ
2.3.11 �Ȱ������ϳ�ø
�Ȱ������ϳ�ø(GS)�����Pֲ�����в���Ȱ������ϳɺ͵���ѭ�����Ȱ������ϳ��Ǹߵ�ֲ�ﵪͬ���ĵ�һ��,��GS����,������Ȱ������������ӵIJ����ĹȰ��������ڴ�л,Ϊ������������������ϳɵ��ṩ����������(glufosinate-ammonium)��GSΪ�б�,ͨ������GS�Ļ���,����Ӳ����ڵ���л���ҺͰ���������,����Ҷ�������,�ƻ��Ӳݹ������,�ﵽ����Ч����
����ѡȡ������Z. mays(PDB��ţ�2D3A,�ֱ��ʣ�2.63 Å)���ഺ˫��˾�Bifidobacterium adolescentis(PDB��ţ�4S17,�ֱ��ʣ�2.30 Å)����˷�֦�˾�Mycobacterium tuberculosis(PDB��ţ�1HTO,�ֱ��ʣ�2.40 Å)����H. sapiens(PDB��ţ�2OJW,�ֱ��ʣ�2.05 Å)��GSA���������ṹ����ͼ(ͼ10)������pymol�����Խ�˷�֦�˾�����(��ɫ)Ϊ������е��Ϻ���,����4�������ϳ̶Ȳ����Ǻܺ�,һЩ�ṹ��Ҳû�������ԡ�
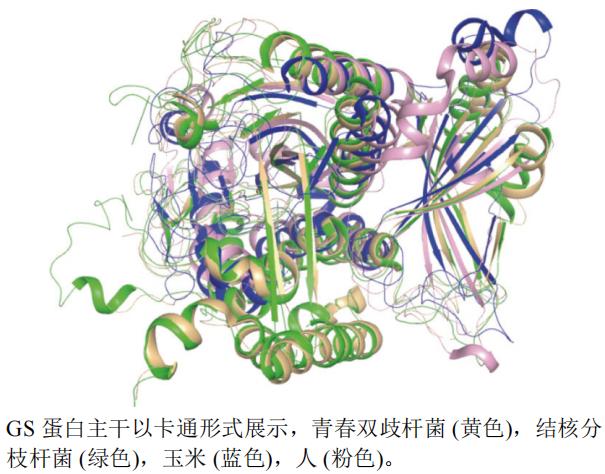
ͼ10 GS��ͬ����A����ά����ͼ
2.3.12 ������֬�����ø
������֬�����ø�ڴ�л;������ϳ����������غ�ľ˨�ء���������(pyroxasulfone)��������֬����ϳ�ø���Ƽ����ݼ�,��Ҫ��Ϊѿǰ��մ�����,�Է�ɱ�̱���Ϊ������Ҷ��Ϊ��,����������ơ��������ﷶΧ��,��������С�����ס�������ˮ�������������
2.4 ��ά�غϳ����
2.4.1 ����װ
ֲ���ܾ�����Ҫ������ѧ����,��ά��ֲ��ϸ����̬�ṹ������ϸ���˶���ϸ�����������䡢�źŴ�����ϸ�����ѵ�,�ɲ����γɷĴ��塢����������������ͻ���ܡ���ë����ë�Ƚṹ����������ݼ���ͨ��������ϸ���ܶ�Ӱ��Ĵ����γ�,������ϸ����˿���ѡ���������������ݼ��������ܵĵ��ʹ���,�����ܵ���ϲ�������ۺ�,��ɷĴ�����ɥʧ,ʹ�Ӳ�ϸ����˿����ͣ����ǰ�ڻ�����,�����쳣�Ķ��κˡ������Ӳ�ϸ������ɥʧ,Һ���γ�������ǿ,�����쳤�����з���������,�����ɸ������͡�
2.4.2 ����֯
����֯���������ϸ�����γ��ܵĽṹ,��������Ҫ�Ĺ��ܣ�1)��֯���ϸ���б�ë����ë���γɣ�2)��֯���ϸ���������ѻ���˿���ѹ����зĴ�����γɡ���������֯�ij��ݼ�ͨ��Ϊ������������,��������(barban)���ȱ�����(chlorpropham),����ͨ���ƻ��Ӳ��ܵ��ȶ������ƻ���˿���Ѷ��ﵽ�������á�
2.4.3 ��ά�غϳ�
ֲ����ά����ά�������Ӫ�����ʽ�����ɵ�˿״����״��,��ֲ�����֧�š����ӡ������ͳ�������á���ື��ݰ�(indaziflam)Ϊ������ά�غϳɵij��ݼ�,�����Ƹ�������ϸ������,�����������ͺͷ����ٻ����������͡�
2.5 �������Ƽ�
2.5.1 ������ģ����
ֲ�����������ɾ��з��Ѻ�������Ե�ϸ������������ֲ�������ٶȺͷ���ļ��ء�������ʹֲ��ϸ�����ɳ�,�Ӷ�ʹϸ�������쳤,������ֲ���л������Ӻ��Ǻ���͵����ʵĺϳɡ��������ص�������,����ģ�����ij�����Ӳݾ���ɱ�����á���2,4-��(2,4-D),����һ��ѡ���Գ��ݼ�,��Ҷ���ݼ�,�ڲ�Ӱ�쵥��Ҷֲ��������ɱ��˫��Ҷֲ��,���ڷ���ˮƽ��ģ����Ȼ�����ء�
2.5.2 ������ת�����Ƽ�
������ת����ֲ�������ͷ��������������,��������Ӱ��Ͳ��������TIBA(2,3,5-triiodobenzoic acid)��NPA(naphthylphthalamic acid)��������õ�������ת�˳��ݼ�,�����Ӳ������������谭�����ص�ת��,����ɱ���Ӳݡ�
2.5.3 ����-D-ľͪ��-5-�����ø
����-D-ľͪ��-5-����ϳ�ø(DXS)�Ǽ�������(MEP);���ĵ�һ������ø,Ҳ�Ǹ�;���Ĺؼ�����ø,�Ե��ƺ�˫���������������Լ�����ܲ��ء�Ҷ���ص���Ҫ���ʵĺϳɾ��йؼ��������á�����DXS�ij��ݼ��ж�����f��ͪ(bixlozone)����f��ͪ(clomazone)��
����ѡȡ�˴˾�E. coli(PDB��ţ�2O1S,�ֱ��ʣ�2.40 Å)���ͷ��������Deinococcus radiodurans(PDB��ţ�2O1X,�ֱ��ʣ�2.90 Å)�����Ͻ�A. thaliana(PDB��ţ�7BZX,�ֱ��ʣ�4.00 Å)��DXSA���������ṹ����ͼ(ͼ11)������pymol���������Ͻ�(��ɫ)Ϊ������Ϻ���,���ߵ��ṹ����,����һ�����ײ����ơ�
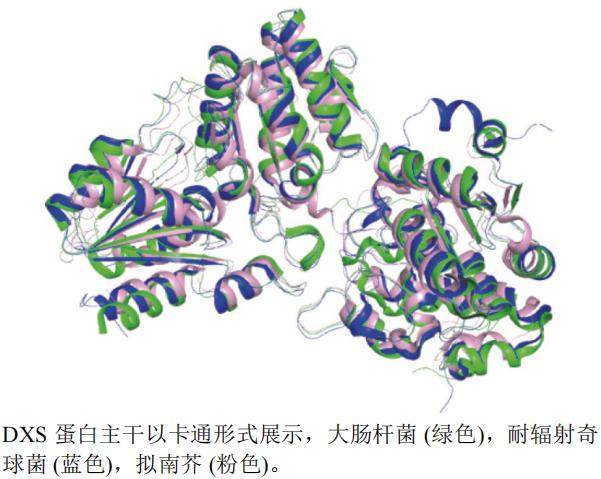
ͼ11 DXS��ͬ����A����ά����ͼ
2.6 ����
2.6.1 ��������������ø
��ऴ�ͷ����ϳ���6��ø�ٲ������,���е�4���ɶ�������������ø(DHODH)������ø�������鵼�Ķ�������������Ϊ������,�����Ӳ�DHODH����λ����������Ĥ�����Ļ��ص���,���ں�����ĺ�������,��;�������ƶԴ�������ﶼ�������ġ�Ŀǰ������ͨ�û�ѧ���Ʊ���ʱ��Ϊtetflupyrolimet�ij��ݼ���ˮ������Ӳݿ��ƾ���ѡ���ԡ�tetflupyrolimet����DHODH�ϵķ������λ��,��tetflupyrolimet����������ֲ��û��ή��,���ᷢչ�����صķ����ٻ�����,��������ȱ�������Ĺؼ�����(���)��
����ѡȡ����H. sapiens(PDB��ţ�1D3G,�ֱ��ʣ�1.60 Å)������Rattus rattus(PDB��ţ�1UUM,�ֱ��ʣ�2.30 Å)��������Trypanosoma brucei(PDB��ţ�2B4G,�ֱ��ʣ�1.95 Å)�����������Lactococcus lactis(PDB��ţ�2BX7,�ֱ��ʣ�2.04 Å)��DHODHA���������ṹ����ͼ(ͼ12)������pymol�����Բ�����(��ɫ)Ϊ������е��Ϻ���,��(��ɫ)�ͺ���(��ɫ)�ĵ��ṹ�������������ײ��ֽṹ�����ơ�
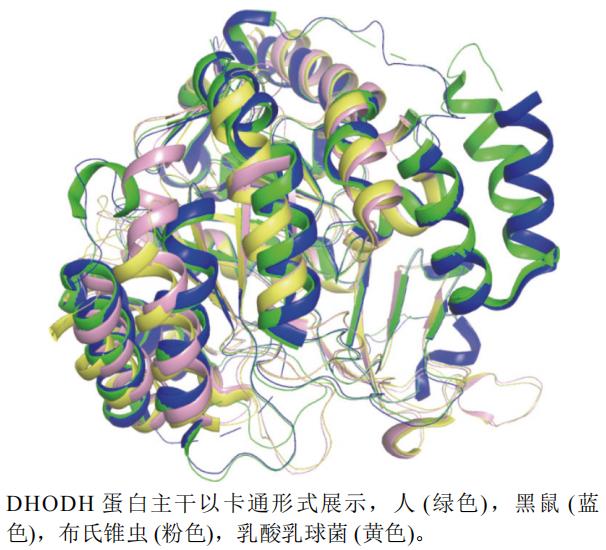
ͼ12 DHODH��ͬ����A����ά����ͼ
2.6.2 �������ϳ�ø
Ҷ����ֲ�����������������Ҫ��������,��������Ҷ�ἰ��������,����ˮ����B��ά����(B9)��Ҷ��ȱ����Ҷ����̬�仯������Ӱ��ֲ����̥���������������������ͽ�ʵ�ȹ��̡�����Ҷ��ij��ݼ��Ʋ���(asulam)��������������˵������з�����β�ݡ����ݡ��[�����ơ��ݵȡ�
����ѡȡ�˴˾�E. coli(PDB��ţ�1AJ0,�ֱ��ʣ�2.00 Å)���������Ⱦ�Thermus thermophilus(PDB��ţ�2DQW,�ֱ��ʣ�1.65 Å)�����������Streptococcus pneumoniae(PDB��ţ�2VEF,�ֱ��ʣ�1.80 Å)������в��˻������Ͼ�Burkholderia cenocepacia(PDB��ţ�2Y5S,�ֱ��ʣ�1.95 Å)�Ķ������ϳ�øA���������ṹ����ͼ(ͼ13)������pymol����������в��˻������Ͼ�(��ɫ)Ϊ������е��Ϻ���,�������������(��ɫ)�ṹ���Կ�ǻ������3�����Ĵ�
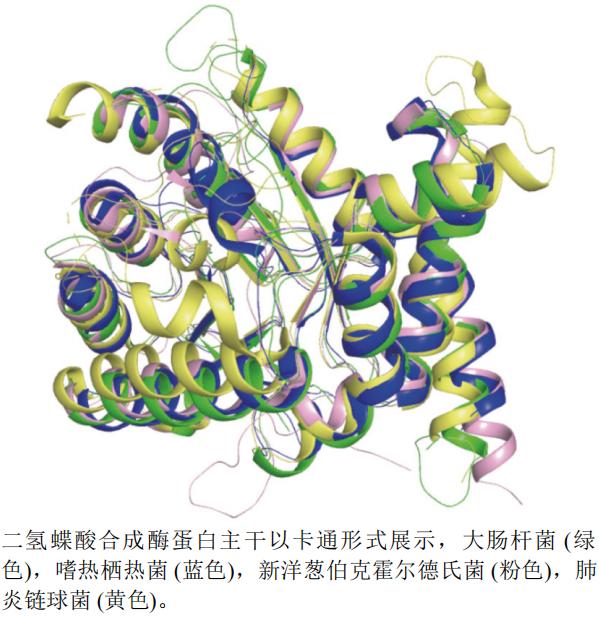
ͼ13 �������ϳ�ø���ײ�ͬ����A����ά����ͼ
3 ���ݼ����Ӱб귢�ֵ���״��չ��
3.1 ���ݼ����Ӱб귢�ֵ���״
Ŀǰ���ݼ����Ӱб귢�����ٺܶ����⣺1)�ݸ�좺Ϳ��ݸ�������Эͬʹ���ںܴ�̶��ϼ����˳��ݼ��б귢�ֵ���������2)���ݼ�������˾�ĺϲ������ٵ�,Ҳ���´��´����о��Ŀ�ѧ�������������,�������ٳ��ݼ��б귢�֡�3)���Ż���Ҫ�������Լ���ũҩ�Ǽ��ƶȸ����ϸ�,���ͳ��ݼ��б귢�ֵijɱ�Ҳ�������ӡ�һ�����ͱ����ǽ�Щ�꽫�ϳ�ũҩ�����г��ijɱ�����2.86����Ԫ,�³��ݼ��б귢�ֺ���ҩ�����Ѷ�Խ��Խ��
3.2 ���ݼ����Ӱб귢�ֵ��¼�����չ��
���ݼ��·��Ӱб�ķ�����Ŀǰ������ݼ�����������÷���֮һ��������ܼ��ַ����·��Ӱб�ķ�����һ���Ի�����ѧΪ�����������\��,Duke��ͨ�����������ر���,����������������������ϳ�;�������ø��������ʽ�Ļ���,֤���˻����鷽��Ϊ���־�����ģʽ���õij��ݼ��ṩ��һ���µIJ��ԡ������˹����ܼ���,���ڼ����ɸѡ�ʹ��ͷ������ݿ�Ĺ���,����ѡ����Ӱб�,ȷ���п��е��³��ݼ����Ӱбꡣ���Ƕ�ѧ���ƽ���չ,��Sukhoverkov����֤������Ҷ����ķ���,����Ϊ�³��ݼ��бꡣHall�ȷ����ڰ���������ϳ�������δ��ҵ��������DZ�ڳ��ݼ��бꡣ��Щ��,��һЩ�����÷�ʽ�ij��ݼ������֣�HMGR(3-�ǻ�-3-���������øA��ԭø)����������������;���ϵ�����ø,������������ߵ��̴�Ѫ֢,Haywood��֤��,������HMGR�������ص�,ʹ��������Ϊ���ݼ��бꣻ��������������ѧ�Ӳ��о�ʵ���ұ����˹��ڵ��²��좽Ӵ����Ե����ص��¼���,���ֲ���������������(ROS)���١���������,����ϸ��Ĥ������֬�ʹ�������ϸ����������,��Ч������ݼ������ճ����ȡ���Щ�����������ǿ�����ԭ�еİб��Ͽ����³��ݼ�,������ֲ����������л;����ȥѰ���µij��ݼ��б�λ�㡣
���������˽�Щ����ݼ��б���������ܡ����ݼ�������ԭ���Լ����÷�Χ,�ڴ˻�����������³��ݼ��б귢�ֵ�����,Ҳ�����˽�Щ��һЩ�����÷�ʽ�ij��ݼ��Ͱб귢�ֵ��¼���,��Ϊ���������µij��ݼ��б��ṩ�ο���